Даже крупнейшие ООПТ — не «чудом сохранившиеся» уголки девственной природы
Поиск по сайту www.EAST-ECO.com


Экологический риск заповедания и пути его преодоления: обзор современных представлений
АВТОР: WOLF_KITSES – 06.06.2018
РУБРИКИ: ЕСТЕСТВОЗНАНИЕ, ЭКОЛОГИЯ И КАПИТАЛИЗМ
Print Friendly Version of this pagePrint Get a PDF version of this webpagePDF
В продолжение темы единой стратегии охраны природы и природопользования
Н.А. Торопова, О.В. Смирнова
Резюме. Даже крупнейшие ООПТ на староосвоенных территориях — не «чудом сохранившиеся» уголки девственной природы, эти ландшафты в предшествующее тысячелетие-два сильно изменены интенсивным природопользованием — подсекой, перелогом и т. д. «кочевыми» формами ведения с/х. Сейчас выясняется, что это же верно для, казалось бы, бесспорно «девственных» участков, вроде хвойных и широколиственных лесов Дальнего Запада США, влажных тропических лесов Амазонии и пр.. Взятие подробных мест под охрану запускает неконтролируемые изменения растительности, почв, влажностного режима, за ними — фауны и пр., при которых виды «теряются» не меньше, чем от природопользования, только иные. Возможны даже гротескные ситуации, когда краснокнижный вид «ликвидируется» заповедным режимом. Отсюда одной охраной недостаточно, чтобы а) сохранить биоразнообразие (это программа-минимум) и б) увеличить его до потенциально возможного, бывшего в доагрикультурный период (программа-максимум), биогеоценозами таких территорий надо управлять, устраивая контролируемые нарушения, подселяя исчезнувшие виды с «вершин» экологической пирамиды, и нижеследующая статья объясняет, как.
***
Потребности общества обусловили необходимость изменить систему организации и управления заповедниками как эталонными объектами, предназначенными для полной реализации экосистемных функций: оптимизации климата и гидрологического режима; поддержания максимального биоразнообразия и продуктивности. Пересмотр представлений теоретической экологии с позиций популяционной парадигмы позволяет выявить причины неполной реализации этих функций в заповедниках и предложить новую методологию и систему мер по восстановлению их природного состояния.
Необходимость оценки экологического риска заповедания
Содержание [скрыть]
1 Необходимость оценки экологического риска заповедания
2 Концепции современной теоретической экологии
2.1 Представления о потенциях и позициях систем
2.2 Элементы популяционной биологии
3 Потенциальный покров
4 Способы решения проблем восстановления и/или поддержания экосистемных функций живого покрова
5 Заключение
На протяжении длительного времени природные экосистемы играли главную роль в реализации экосистемных функций [1–7]. Однако катастрофическое сокращение их площадей в результате многовекового природопользования привели к неспособности большей части экосистем полностью реализовать свои функции [3–7].
В мировой литературе накоплены данные о роли природопользования в формировании современного облика экосистем. На основе дендрохронологических, палинологических, исторических данных и радиоуглеродного анализа углей прослежены этапы антропогенной деградации природных экосистем с конца палеолита – неолита до современности [1; 7–11].
Исследования в заповедниках Европейской России выявили существенный экологический риск заповедания лесных и лесостепных территорий. Показано, что режим полной охраны ведет к сокращению видового разнообразия, исчезновению лугово-степных сообществ и лесов со светолюбивыми видами, изменению локального климата и гидрологического режима [9; 11; 12]. Основная причина – предшествующее преобразование этих территорий в циклах традиционного природопользования: подсечно-огневого и пахотного земледелия, выпаса скота, сбора подстилки, рубок и пр. В результате этих воздействий полностью уничтожены ключевые виды животных лесных и лесостепных ландшафтов (зубры, туры, тарпаны, бобры и пр.) и существенно изменены ареалы видов деревьев, определявших организацию природного покрова. Человек стал определять все параметры этого покрова с начала позднего голоцена, выступая как самый мощный ключевой вид [1–2; 8–9; 11; 13–14].
Следует особо подчеркнуть, что в большинстве случаев изъятие заповедных территорий из хозяйственной деятельности не сопровождалось восстановлением природного разнообразия и не определялись размеры территорий, необходимых для устойчивого существования популяций ключевых видов. Это привело и приводит в настоящее время к падению биоразнообразия и к невозможности в полной мере реализовать остальные экосистемные функции [3; 7; 10; 13; 15–18].
Современное состояние экосистем заповедников настоятельно требует разносторонних исследований механизмов осуществления экосистемных функций, как природных экосистем, так и экосистем на разных стадиях антропогенных дигрессий. Это первый шаг в разработке экологически обоснованной стратегии охраны и природопользования [9; 11–12].
Стации обитания зубров в Кизгичском лесничестве Тебердинского государственного биосферного заповедника: а, б – прирусловые луга; в – опушки; г – прирусловые ивово-сероольховые груды (а–г – Архыз, Шевченко Н. Е., 2014)
Стации обитания зубров в Кизгичском лесничестве Тебердинского государственного биосферного заповедника: а, б – прирусловые луга; в – опушки; г – прирусловые ивово-сероольховые груды (а–г – Архыз, Шевченко Н. Е., 2014)
Концепции современной теоретической экологии
Решение проблемы сохранения и восстановления экосистемных функций основывается на закономерностях организации природных экосистем и их ландшафтных комплексов как эталонных объектов. Теоретические основы их познания сформулированы в результате переосмысления классических концепций экологии экосистем с позиций системного анализа и популяционной парадигмы [8; 16; 19].
Представления о потенциях и позициях систем
Для решения поставленной проблемы принципиально важно разделять представления о потенциях и позициях системы. Потенции системы – это ее свойства, полностью проявляющиеся при спонтанном развитии в оптимальных условиях. Если оно нарушается внешними воздействиями и/или осуществляется в неоптимальных условиях, реализуется только часть свойств, и это характеризует позиции системы в конкретных условиях. Представление о потенциях системы и ее параметрах позволяет сформировать логически непротиворечивую концепцию развития, не нарушаемого извне. Описание развития экосистемы при полной реализации потенций – удобная модель, «эталонный вариант», нужный для оценки степени отклонения от него реальных экосистем и выяснения причин этого.
Представления о потенциях и позициях в равной степени относятся как к отдельной экосистеме, так и к ландшафтным комплексам локального и регионально уровня и к биогеоценотическому покрову в целом. Выявление потенций природных экосистем и их комплексов на основе натурных исследований и моделирования – основа современной синэкологии [8].
Среди экосистем суши наиболее полно исследованы природные закономерности лесных экосистем, отраженные в концепциях: «gap mosaic concept» и «mosaic-cycle concept of ecosystems» [1; 11–12].
Элементы популяционной биологии
Живой (биотический) покров представляет собой множество взаимодействующих популяций видов [8; 11; 20–21] различных трофических групп. С экологических позиций элементарная популяция (ЭП) – это множество особей одного вида, необходимое и достаточное для поддержания устойчивого потока поколений.
ЭП характеризуются: 1) временем жизни одного поколения; 2) минимальным пространством, необходимым для устойчивого потока поколений; 3) экологической плотностью (числом или массой особей в чистой заросли на единицу площади); 4) специфическим размещением популяционных локусов в пространстве [1; 8; 21]. ЭП разных видов и трофических групп по значениям каждого из перечисленных признаков располагаются в континуальные ряды. Виды с наиболее крупными и длительно существующими популяционными мозаиками, включающие в циклы оборота поколений наибольшую порцию вещества и энергии, мы считаем мощными средопреобразователями. Их называют: “keystone species” (ключевые виды) и “ecosystem engineers” (экосистемные инженеры). Исходное понятие – «эдификатор» [8; 12; 21]. Ключевые виды в процессе спонтанного развития наиболее значимо преобразуют как местообитания ЭП в целом, так и местообитания их элементов. Это ведет к изменению гидрологического, температурного, светового режимов; микро-, мезорельефа; строения почвенного покрова и пр.
Внутренняя гетерогенность местообитания ЭП ключевого вида позволяет существовать вместе экологически и биологически различным подчиненным видам и поддерживает высокий уровень биоразнообразия [8; 20–21]. В природных лесных ландшафтах устойчиво поддерживалась мозаичность разного типа, создаваемая наиболее мощными ключевыми видами. Фитогенная мозаичность поддерживалась вследствие образования: 1) прорывов в пологе леса («gaps») в результате старения и естественной смерти одного-нескольких рядом растущих деревьев; 2) ветровально-почвенных комплексов («tree fall»), возникающих в том случае, когда смерть и падение дерева сопровождается пертурбацией и создается специфический «вывальный» микрорельеф [11].
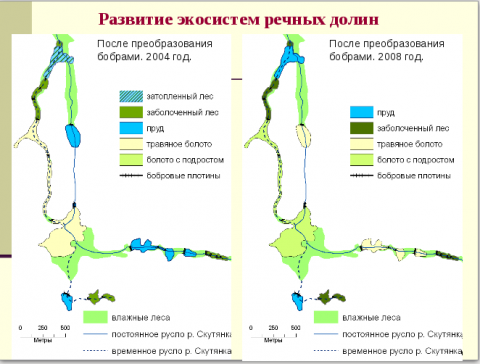
Зоогенную мозаичность поддерживалась в результате: 1) уничтожения крупными копытными подроста и молодых деревьев, кустарников, лесных трав, уплотнения и унавоживания почв и возникновения полян с лугово-опушечной и лугово-степной флорой; 2) уничтожения листо- и хвоегрызущими насекомыми листьев и хвои в кронах взрослых деревьев, увеличения интенсивности солнечной радиации на поверхности почвы, повышение температуры воздуха и почвы, обогащение почвы азотом и другими минеральными веществами из экскрементов, возрастания численности светолюбивых и нитрофильных видов трав; 3) строительства бобрами на ручьях и мелких речках плотин и каналов, формирования прудов и низинных болот, увеличения видового разнообразия и численности сопутствующих видов растений и животных; 4) избирательного уничтожения бобрами деревьев и кустарников по берегам водоемов и развитие прирусловых луговых полян со светолюбивой флорой и фауной [11–12; 21]. Кроме фитогенной и зоогенной мозаичности более мелкие элементы мозаики связаны с деятельностью грибов (микогенная мозаичность) [12; 20].
Снимок экрана от 2018-05-29 23:33:47
Здесь и далее источник — презентации к лекциям О.В.Смирновой
Постоянное участие в доагрикультурных лесах животных-фитофагов создавало четко выраженную гетерогенность среды: собственно лесные участки чередовались с зоогенными полянами и зоогенными водоемами. При этом весь комплекс сообществ лесных ландшафтов мог существовать только на площади, превышающей площадь элементарной популяции ключевого вида с самыми крупными мозаиками, В лесных ландшафтах, где самые крупные мозаики образуют виды зубрового комплекса, – это сотни и более км2 (табл. 1).
Существование на одной и той же территории иерархий мозаик, создаваемых ключевыми видами растений и животных, определяло в доисторический период спонтанное развитие лесолуговых ландшафтов и поддержание максимального видового разнообразия за счет создания максимальной гетерогенности среды.
В природных степных ландшафтах господствующий тип мозаичности – зоогенный. Он формируется вследствие популяционной жизни ключевых видов животных. В ненарушенных евразийских степях и полупустынях и североамериканских прериях – это роющие мелкие позвоночные (сурки, суслики, песчанки, гоферы, луговые собачки и др.) и крупные стадные копытные (лошади, куланы, сайгаки, бизоны и др.) [8; 20–21]. В доагрикультурных степях и полупустынях зоогенная мозаика создается и поддерживается в результате:
1) создания микро- и мезорельефа роющей деятельностью мелких позвоночных, перемещения нижних почвенных горизонтов на дневную поверхность, увеличения физической и химической эрозии подстилающих пород, иссушения и засоления почв и возникновения ксерофитных и галофитных группировок растений;
2) локального уничтожения крупными копытными и мелкими роющими животными растительного покрова, развития процессов эрозии, развеивания легких субстратов и возникновения участков, лишенных растительности;
3) уплотнения и унавоживания почв на выбросах грунта из нор мелких роющих животных и возникновения нитрофильных и рудеральных группировок растений;
4) перераспределения атмосферных осадков, увеличения увлажнения в микро- и мезозападинах, возникших на местах заброшенных нор, рассоления, увеличения содержания гумуса и образования группировок гликофитных и мезофитных видов [21–22].
Размеры и время существования мозаик, создаваемых разными животными, существенно различаются, что определяет наличие разновременных и разномасштабных процессов смен в спонтанных сообществах. В связи с этим, как и в лесном поясе, устойчивое существование степных ландшафтов возможно только на площади, большей площади ЭП ключевых видов с самыми крупными мозаиками. В степных ландшафтах Евразии – это тысячи-десятки тысяч км2 [1; 12; 20–22].
Снимок экрана от 2018-05-29 23:34:38
Потенциальный покров
Представление о ключевых видах – основа реконструкций потенций экосистем и потенциального покрова в целом как эталонного объекта для оценки экосистемных функций. Учитывая невозможность полностью оценить биологическое разнообразие экосистем в целом (включая микромир), под потенциальным покровом на современном уровне знаний мы понимаем такой покров, в котором устойчиво существуют популяции всех сохранившиеся до настоящего времени ключевых видов и сопряженных компаний подчиненных видов, выявленных к настоящему времени [8]. В современном покрове практически нет экосистем и ландшафтных комплексов, полностью соответствующих представлениям о потенциальном покрове. Однако сохранившиеся фрагменты потенциального покрова в заповедниках в разной степени способны к восстановлению в спонтанном режиме или при антропогенном содействии, в первую очередь при условии реинтродукции природных ключевых видов [1; 12; 20].
Способы решения проблем восстановления и/или поддержания экосистемных функций живого покрова
Оценка современного состояния заповедных территорий основана на решении следующих задач:
1) создание модельной реконструкции потенциального покрова на основе исторических, археологических и палеонтологических данных;
2) оценка потерь биологического разнообразия;
3) оценка возможности восстановления популяций ключевых видов сопряженных компаний подчиненных видов в современных экотопических и климатических условиях анализируемой территории;
4) определение размеров территории, необходимой для устойчивого существования ключевых видов, живущих на данной территории или способных устойчиво существовать при условии их естественной или антропогенной реинтродукции;
5) построение сукцессионных систем и расчеты времени восстановления устойчивых потоков поколений ключевых видов.
К настоящему времени накоплен большой опыт по реконструкции потенциального покрова, оценке сукцессионного статуса экосистем и составлению прогнозов развития экосистем и их ландшафтных комплексов, в т. ч. и на заповедных территориях [1; 9; 12; 13; 23; 24]. Эти исследования составляют базовую основу решения задач:
1) ориентация систем природопользования на потенциальные возможности территорий, включая оптимальный гидрологический режим и максимальное биологическое разнообразие;
2) планирование членения территории на локальном, региональном и континентальном уровнях в соответствии с уровнем сохранения биоразнообразия;
3) выделение в качестве заповедных тех территорий, где в спонтанном режиме поддерживается высокий уровень биоразнообразия;
4) разделение существующих ООПТ на комплексы экосистем, способных к устойчивому поддержанию биоразнообразия в связи с сохранившимся комплексом природных ключевых видов (лесные экосистемы) и поддерживаемых только антропогенным путем (луговые и лугово-степные экосистемы);
5) организация природопользования с учетом сохранения максимально возможного природного биоразнообразия (замена монокультур в лесоводстве и сельском хозяйстве поликультурами, культивирование природных видов);
6) определение размеров хозяйственных секций на основе учета дальности миграций природных видов (радиус распространения зачатков природной флоры и дальности расселения животных);
7) размещение хозяйственных секций в пределах природных угодий (лесов, луговых, степных, пустынных и тундровых пастбищ), имитирующее природную мозаичность;
8) организация резерватов природных видов на местах крупных разрывов ареалов ключевых видов вследствие антропогенных преобразований биот (лесные резерваты с восстановлением природного разнообразия, восстановления участков степной и луговой растительности и пр.);
9) организация регионального мониторинга ключевых видов.
Снимок экрана от 2018-05-29 23:35:31
Заключение
Обзор теоретических представлений и практических разработок современной системной экологии показывает реальную возможность и конкретные пути предотвращения экологического риска заповедания, которое было проведено без достаточного учета уровня деградации биогеоценотического покрова. Опыты по реинтродукции ключевых видов животных и растений, проводимые в нашей стране и других странах на протяжении многих десятилетий, свидетельствуют об успешности этих усилий. Однако современное состояние биогеоценотического покрова требует решения проблем экосистемных функций в глобальном масштабе.
Для этого в нашей стране необходимо централизованное сотрудничество вузовской и академической науки с сотрудниками ООПТ, специалистами лесного хозяйства и управляющими органами.
Снимок экрана от 2018-05-29 23:34:22
Литература
1. Мониторинг биоразнообразия лесов: методология и методы / отв. ред. А.С. Исаев. М.: Наука, 2008. 453 с.
2. Павлов Д.С., Букварева Е.Н. Климаторегулирующие функции наземных экосистем и экологоцентрическая концепция природопользования // Успехи современной биологии. 2011. Т. 131. No 4. С. 324-345.
3. Gamfeldt L., Hillebrand H., Jonsson P.R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning // Ecology. 2008. V. 89 (5). Р. 122-1231.
4. Hector, A., Bagchi R. Biodiversity and ecosystem multifunctionality // Nature. 2007. V. 448. Р. 188-190.
5. Hooper D.U., Chapin F.S., Ewel J.J., Hector A., Inchausti P., Lavorel S., Lawton J.H., Lodge D.M., Loreau M., Naeem S., Schmid B., Setala H., Symstad A.J., Vandermeer J., Wardlw D.A. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge // Ecol. Monogr. 2005. Т. 75. No 1. С. 3-35.
6. Mysterud A., Barton K.A., Jedrzejewska B., Krasinski Z.A., Niedziałkowska M., Kamler J.F., Naaf T., Wulf M. Effects of gap size, light and herbivory on the herb layer vegetation in European beech forest gaps // Forest ecology and management. 2007. V. 244. P. 141-149. 7.7. Chiarucci A,. Arau ́jo M.B., Decocq G., Beierkuhnlein C., Ferna ́ndez- 7/
Palacios J.M., Ciccarese L., Mattsson A., Pettenella D. Ecosystem services from forest restoration: thinking ahead // New For. 2012. Т. 43. No 5-6. С. 543-560.
8. Смирнова О.В., Торопова Н.А. Сукцессия и климакс как экосистемный процесс // Успехи совр. биол. 2008. No 2. С. 129-144.
9. Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России / отв. ред. Л.Б. Заугольнова. М.: Научный мир, 2000. 185 с.
10. Bobiec A., Burgt H., Meijer K., Zuyderduyn C., Haga J., Vlaanderen B. Rich deciduous forests in Białowieża as a dynamic mosaic of developmental phases: premises for nature conservation and restoration management // Forest ecology and management. 2000. V. 130. P. 159-175.
11. Бобровский М.В. Лесные почвы европейской России: биотические и антропогенные факторы формирования. М.: КМК, 2010. 359 с.
12. Восточноевропейские леса: история в голоцене и современность / отв. ред. О.В. Смирнова. М.: Наука, 2004. Кн. 1. 479 с. Кн. 2. 575 с.
13. Чумаченко С.И., Смирнова О.В. Моделирование сукцессионной динамики насаждений // Лесоведение. 2009. No 6. С. 3-17.
14. Vera F.W.M., Bakker E., Olff H. Large herbivores: missing partners of western European light-demanding tree and shrub species? // Large herbivore ecology, ecosystem dynamics and conservation / еd. by K. Danell, P. Duncan, R. Bergstrom, J. Pastor. Biology series. Cam-
bridge University Press, 2006. P. 203-231.
15. Schwartz M.W., Brigham C.A.,· Hoeksema J.D., LyonsK.G., Mills M.H.,· Mantgem P.J. Linking biodiversity to ecosystem function: implications for conservation ecology // Oecologia. 2000. V. 122. Р. 297-305.
16. Смирнова О.В. Методология исследования экосистем с популяционных позиций // Известия ПГПУ им. В.Г. Белинского. 2011. No 25. С. 15-21.
17. Prach K., Sándor B., Joyce C.B.; Pyšek P., van Diggelen R., Wiegleb G. The role of spontaneous vegetation succession in ecosystem restoration: A perspective // Applied Vegetation Science. 2001. V. 4. Р. 111-114.
18. Schwartz M.W., Brigham C.A.,· Hoeksema J.D., Lyons K.G., Mills M.H.,· Mantgem P.J. Linking biodiversity to ecosystem function: implications for conservation ecology // Oecologia. 2000. V. 122. Р. 297- 305.
19. Taylor A.R., Chen H.Y.H., VanDamme L. A review of forest succession models and their suitability for forest management planning // Forest science. 2009. V. 55 (1). Р. 23-36.
20. Смирнова О.В. Методологические подходы и методы оценки климаксового и сукцессионного состояния лесных экосистем (на примере восточноевропейских лесов) // Лесоведение. 2004. No 3. С. 15-26.
21. Методические подходы к экологической оценке лесного покрова бассейна малой реки / отв. ред. Л.Б. Заугольнова, Т.Ю. Браславская. М.: КМК, 2010. 383 с.
22. Смирнова О.В. Популяционная организация биоценотического покрова лесных ландшафтов // Успехи совр. биологии. 1998. No 2. С. 25-39.
23. Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия / под ред. О.В. Смирновой, Е.С. Шапошникова. СПб.: РБО, 1999. 549 с.
24. Смирнова О.В. Теоретические основы, система методов и программ стационарных экологических исследований в таежных лесах Урала // Труды Печоро-Илычского заповедника. Сыктывкар, 2010. Вып. 16. С. 157-162.
Вестник ТГУ, т.19, вып.5, 2014
http://www.socialcompas.com/2018/06/06/ekologicheskij-risk-zapovedaniya-...




- Войдите чтобы оставить комментарии
Сайт существует на частные пожертвования. Мы будем рады любой поддержке.
Нам важно знать, что вы нас цените. --> ПОДДЕРЖАТЬ
